本網(wǎng)訊茶毫是一種茶葉表皮毛,呈透明細(xì)長(zhǎng)卷絲狀�����,頂端無(wú)分叉,主要分布在幼嫩芽葉的背部��,并隨著幼葉成熟而自行脫落�����。茶毫與擬南芥表皮毛類(lèi)似,都屬于單細(xì)胞類(lèi)型表皮毛����。茶毫是茶樹(shù)重要的農(nóng)藝性狀,含有豐富多樣的次生代謝物�����,對(duì)茶葉品質(zhì)形成非常重要�����。多種類(lèi)型的茶葉加工工藝都單獨(dú)設(shè)置“提毫”流程��,如名優(yōu)綠茶碧螺春����、黃山毛峰等以“白毫顯露”為優(yōu)����;高級(jí)祁紅、滇紅以“金毫顯露”為優(yōu)�����;白毫銀針、白牡丹以“銀毫顯露”為優(yōu)等�。此外,茶毫對(duì)茶樹(shù)抵御外界病蟲(chóng)害���、紫外脅迫等也具有重要作用����。然而茶毫發(fā)育的調(diào)控機(jī)制目前還尚不清楚����。
近日,我校茶樹(shù)生物學(xué)與資源利用國(guó)家重點(diǎn)實(shí)驗(yàn)室趙劍課題組在國(guó)際期刊New Phytologist上發(fā)表了題為“CsMYB1 integrates the regulation of trichome development and catechins biosynthesis in tea plant domestication”的研究論文�,揭示了MYB轉(zhuǎn)錄因子CsMYB1協(xié)同調(diào)控茶毫發(fā)育和兒茶素合成,及在茶樹(shù)從野生到栽培馴化過(guò)程中的分子遺傳變異機(jī)制�����。

該研究對(duì)茶樹(shù)種質(zhì)資源茶毫表型進(jìn)行了調(diào)查��。發(fā)現(xiàn)栽培種茶樹(shù)的幼嫩葉片一般包被細(xì)長(zhǎng)茶毫����,而大部分茶樹(shù)野生近源種的茶毫相對(duì)較少����。對(duì)60個(gè)栽培茶樹(shù)基因表達(dá)和茶毫表型關(guān)聯(lián)分析�����,結(jié)合進(jìn)化樹(shù)分析等手段篩選出一個(gè)與茶毫表型關(guān)系密切的候選基因CsMYB1���,該基因是擬南芥GL1的同源基因���,主要表達(dá)在茶毫富集的幼嫩組織中��,如芽�����,第一葉等�����。進(jìn)一步結(jié)果表明��,CsMYB1完全互補(bǔ)atgl1突變體表皮毛缺陷表型���,CsMYB1上調(diào)AtGL2等下游基因表達(dá)���。AsODN和茶樹(shù)轉(zhuǎn)基因發(fā)根實(shí)驗(yàn)也表明���,CsMYB1直接調(diào)控茶樹(shù)GL2、CPC���、GL3等基因表達(dá)�����。酵母雙雜交��、BIFC��、EMSA�����、啟動(dòng)子激活等實(shí)驗(yàn)結(jié)果表明���,CsMYB1與CsGL3,CsWD40等蛋白互作����,形成MBW(MYB-bHLH-WD40)復(fù)合體調(diào)控GL2基因表達(dá)���,而CsCPC則與CsMYB1競(jìng)爭(zhēng)性地與CsGL3,CsWD40互作���,負(fù)調(diào)控GL2基因表達(dá)�����。生物信息學(xué)分析發(fā)現(xiàn)CsMYB1的表達(dá)模式與兒茶素含量分布以及相關(guān)合成酶基因表達(dá)模式高度相關(guān)��。在逆境脅迫下����,CsMYB1的表達(dá)模式與兒茶素的含量變異也呈現(xiàn)顯著正相關(guān)�。通過(guò)AsODN和茶樹(shù)轉(zhuǎn)基因發(fā)根實(shí)驗(yàn)表明��,CsMYB1能直接結(jié)合到ANR等下游基因啟動(dòng)子區(qū)���,以MBW復(fù)合體的形式調(diào)控下游靶基因的表達(dá)��,調(diào)控茶樹(shù)兒茶素的合成(Figure 1)�。
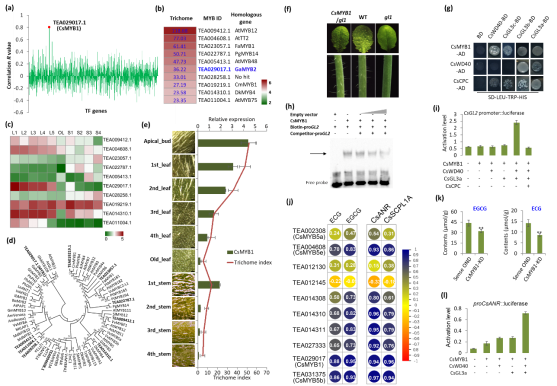
Figure 1:CsMYB1調(diào)控茶毫發(fā)育和兒茶素類(lèi)化合物合成的分子機(jī)制
此外,大部分野生茶樹(shù)資源的茶毫指數(shù)顯著低于栽培品種����,表現(xiàn)出明顯的選擇/馴化特征。本研究表明茶樹(shù)從野生到栽培進(jìn)化過(guò)程中茶毫的選擇/馴化特征與CsMYB1基因變異密切有關(guān)����。群體遺傳證據(jù)表明茶樹(shù)從野生種到古茶樹(shù)再到現(xiàn)代栽培種進(jìn)化過(guò)程中,CsMYB1的遺傳多樣性呈現(xiàn)明顯下降趨勢(shì)�,這暗示了該基因在茶樹(shù)進(jìn)化過(guò)程中可能受到了自然或人工選擇。對(duì)CsMYB1啟動(dòng)子序列的分析發(fā)現(xiàn)�����,與野生茶樹(shù)相比�,栽培茶樹(shù)中CsMYB1啟動(dòng)子有192bp的序列插入,該序列中包含多個(gè)轉(zhuǎn)錄增強(qiáng)子���,并且與茶毫指數(shù)����、兒茶素含量���,以及CsMYB1表達(dá)水平都顯著相關(guān)��。啟動(dòng)子GUS酶活實(shí)驗(yàn)進(jìn)一步表明�,栽培茶樹(shù)CsMYB1啟動(dòng)子(含有192bp插入)相比野生茶樹(shù)具有更高的轉(zhuǎn)錄活性(Figure 2)。
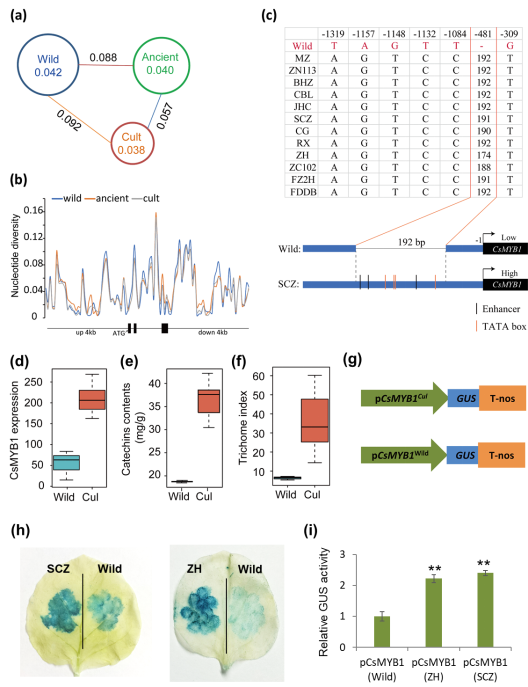
Figure 2:茶樹(shù)群體中CsMYB1遺傳變異機(jī)制
本研究揭示了CsMYB1在茶毫發(fā)育和兒茶素合成調(diào)控方面的雙重功能 (Figure 3)��,初步明確了該基因在茶樹(shù)馴化過(guò)程中的作用機(jī)制�,為培育優(yōu)質(zhì)高抗茶樹(shù)品種提供了理論依據(jù)。
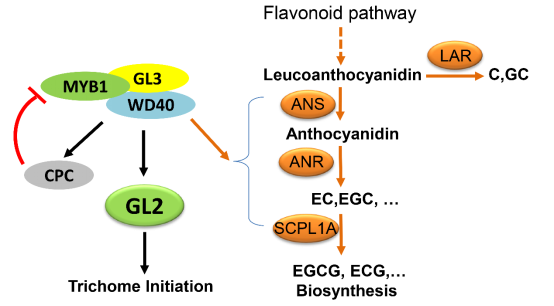
Figure 3:茶樹(shù)中CsMYB1調(diào)控機(jī)制模型
我校茶與食品科技學(xué)院李鵬輝副教授為第一作者����,趙劍教授為通訊作者。童偉副教授��、夏恩華教授以及臨沂大學(xué)王樹(shù)才教授等參與了研究�。此研究受?chē)?guó)家自然科學(xué)基金、國(guó)家重點(diǎn)研發(fā)計(jì)劃�、安徽農(nóng)業(yè)大學(xué)神農(nóng)學(xué)者人才計(jì)劃等資助。
文章鏈接:https://nph.onlinelibrary.wiley.com/doi/abs/10.1111/nph.18026





















